Ferritin
.png)
Iron is an essential element for living organisms. It has crucial roles in processes such as oxygen transport and cellular energy production, and can also serve as a cofactor or catalyst in enzymatic reactions (Arosio et al., 2017). Nonetheless, excess iron might lead to adverse reactions, including toxicity and oxidative stress. The cell possesses an intricate system whose function is to balance cellular iron import, export, and storage to maintain iron homeostasis. Such a system involvesferritin, an iron storage protein.
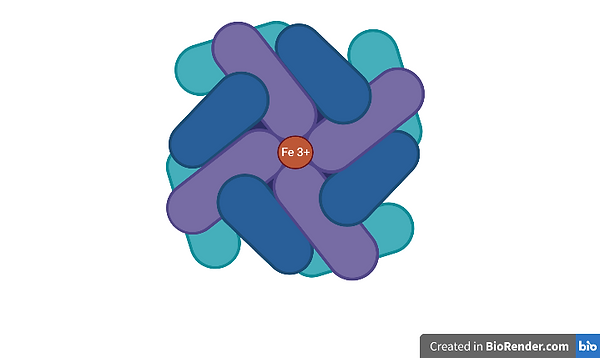
Ferritin is a multimeric protein that forms a nanocage structure that can contain up to around 4500–5000 iron atoms. In humans, ferritin has a mass of 474 kDa and a total of 24 subunits. It is ubiquitous in all forms of cellular life, with the exception of yeast, and has wide-ranging functions other than iron storage. Ferritin makes iron available for critical cellular processes while protecting lipids, DNA, and proteins from the potentially toxic effects of iron (Bellini et al., 2014). Changes in the structure of ferritin are frequently seen in clinical practise and are often shown through defects in iron homeostasis or metabolism. Its primary role is in iron sequestration, where it functions as a ferroxidase, thus converting Fe(II) to Fe(III) as iron is internalised and sequestered in the ferritin mineral core (Bellini et al., 2014).
.png)
Figure 1 - Structure of Ferritin
Ferritin is also found extracellularly within the serum, where it serves as an important clinical marker of iron status. It is regullary used in clinical medicine but the precise source of serum ferritin has yet to be determined (Knovich et al., 2008).
Ferritin plays a role in ferroptosis, an iron-dependent form of cell death. Ferroptosis results from the accumulation of iron‐dependent lipid peroxide, and differs from apoptosis due to the mechanism underlying ferroptosis induction. This form of programmed cell death was found to have important implications in conditions and diseases such as Alzheimer's disease (AD), cancer, COVID-19, hyperferritinemia and kidney disease. It is becoming more recognised that ferritin also plays a role in a multitude of other conditions, such as inflammatory, neurodegenerative, and malignant diseases. Ferritin has a significant use as a diagnostic and prognostic marker and also has other applications in bionanotechnology.
.png)
Figure 2 - Ferroptosis Pathway
Hereditary Hyperferritinemia
An uncommon condition known as hereditary hyperferritinemia-cataract syndrome (HHCS) is characterised by elevated serum ferritin levels, cloudy cataracts, and a lack of iron storage in tissues. This disorder originated from mutations in the untranslated regions of the light ferritin gene. This causes a surplus of ferritin, which accumulates in the serum and precipitates in the lens of the eye. This causes a condition known as cataracts (Celma Nos et al., 2021).
Two main theories have been put forth to explain why cataracts arise in HHCS. The first theoryrelates to iron homeostasis and excessive L-ferritin production could lead to an increase in free iron and reactive oxygen species. This leads to oxidative lens damage that results in a loss of transparency. This theory, however, is refuted by the iron-poor nature of the lens-diffracting crystals and the fact that L-Ferritin does not directly bind iron.
The second and most widely accepted theory states that excessive crystal deposits made of aggregated and iron-poor L-ferritin cause the loss of appropriate lens transparency (Celma Nos et al., 2021). Other than bilateral cataracts, the HHCS patients don't exhibit any other clinical signs. When individuals with HHCS experience visual impairment due to the development of cataracts, typically in the adult stage, they are treated by having surgical surgery (Celma Nos et al., 2021).

Image 3 - Hereditary Hyperferritinemia Mechanism Pathway
Ferritin & Restless Leg Syndrome
Restless Leg Syndrome (RLS) is a neuronal movement disorder that frequently occurs in tandem with sleep problems. RLS patients frequently experience uncomfortable sensations that are worse during periods of inactivity and frequently interfere with sleep, and cause them to feel compelled to move their legs. Interestingly, 33.3% of pregnant women with RLS reported symptoms prior to becoming pregnant, thus 66.7% of women experienced symptoms during pregnancy, and of those women, 97% of those affected reported that their symptoms diminished within 5 days of giving birth (Schulte et al., 2014). Serum ferritin levels were lowered in 83% of the 24 patients with restless legs syndrome in a study conducted by Kotagal and Silber (2004), indicating that iron insufficiency is a defining feature of the condition.
Restless leg syndrome is also associated with iron deficiency. A patient's iron reserves could be inadequate even without observable anaemia. Recent research has demonstrated that reduced iron stores, which are noted by serum ferritin levels below 50 ng/mL, might aggravate RLS symptoms. Serum ferritin levels should be checked in patients with recently diagnosed RLS or in RLS patients whose symptoms have recently worsened (THORPY et al., 2000).
There has been some speculation that patients with restless legs syndrome have much lower spinal fluid ferritin levels than controls (individuals not diagnosed with RLS), suggesting a cause for the illness. The neuromelanin cells may have trouble absorbing iron, which can lead to restless legs syndrome. There are still many RLS patients with low blood ferritin levels who appear to benefit from iron therapy in the current therapeutic environment (Connor et al., 2003).
However, the situation is not as straightforward as one might think. It has been found that other iron pathway components are altered in RLS patients. Genetic factors associated with RLS suggest an influence on iron metabolism that has not yet been fully explored. Moreover, people who have hemochromatosis-induced systemic iron overload are not immune to RLS (Schulte et al., 2014).
.png)
Image 4 - Role of Ferritin in Restless Leg Syndrome
Ferritin & Covid-19
Medical professionals and researchers were both intrigued by elevated ferritin levels amongst COVID-19 patients. Unfavourable results were anticipated both before and immediately after the COVID-19 pandemic's global spread. However, the possible association between ferritin and a worse prognosis is only a small portion of the hidden elements underlying it; hence, more investigation is needed. Ferritin levels then advanced from being a prognostic factor indicator to a risk factor for severity (Alkattan et al., 2021; Chen et al., 2020). Moreover, a sudden increase in ferritin levels is frequently accompanied by coagulopathies, decreased NK cell quantity and activity, abnormal liver function tests, and lymphopenia. This led researchers to hypothesise and later affirm that COVID-19 might be the newest addition to the family of hyperferritinemic syndromes. COVID-19 symptoms that are similar to hyperferritinemic syndromes include systemic inflammatory response syndromes (SIRS) and acute respiratory disease syndromes (ARDS). A common cause of hyperferritinemic syndrome and its complications is a combination of high serum ferritin levels and life-threatening hyperinflammation, which can eventually lead to multi-organ failure.
.png)
Image 5 - Role of Ferritin in Covid19
Ferritin has also been applied in the production of vaccines. One example is the COVID-19 vaccine, which uses ferritin nanoparticles to create a spike protein receptor-binding domain vaccine. The COVID-19 vaccine was shown to induce an effective antibody immune response in mice that persisted for around 7 months while also maintaining a high population of memory B cells (Wang et al., 2021).
Ferritin & Chronic Liver Disease
Haematological abnormalities and chronic liver diseases usually coexist. Patients with liver cirrhosis usually show symptoms of iron overload or of iron deficiency anaemia. The symptoms of iron overload come from a decrease in the function of the iron entry regulator protein hepcidin. This decrease in function is due to a decrease in the liver's synthetic function. On the other hand, iron deficiency in chronic liver illnesses, may be caused by bleeding as a result of portal hypertension. Patients with severe late-stage chronic liver disease exhibit inflammation as well, and as a result, such chronic liver illnesses are frequently associated with elevated hepatic and serum ferritin levels (Cotler et al., 1998; Eng et al., 2005). Ferritin levels in the serum may act as a prognostic biomarker in patients suffering from liver disease due to the correlation between their concentrations and hepatic histological lesions in patients suffering from non-hemochromatosis liver illnesses. These include end stage liver diseases characterised by an accumulation of iron in the liver. A 2017 study tracked the serum ferritin levels of patients with decompensated cirrhosis for 6 years and discovered that high levels of ferritin are related to a more catastrophic clinical outcome (Oikonomou et al., 2017).
.png)
Image 6 - Role of Ferritin in Chronic Liver Disease
Ferritin as a Diagnostic Tool
Ferritin can also be described as an invaluable tool in the clinical setting , as it possesses a wide variety of diagnostic uses that can range from the identification of common diseases such as iron deficiency anaemia (IDA) to the diagnosis of hereditary or acquired iron overload diseases such as hereditary hemochromatosis and chronic transfusion therapy (Theil, 2012).
Iron is stored intracellularly as ferritin, which is an acute-phase reactant. The increase in ferritin synthesis acts as a nonspecific response that is part of the general pattern of the systemic effects of inflammation. Fever, acute infections, rheumatoid arthritis, and other chronic inflammatory disorders elevate the plasma ferritin concentration. Therefore, the diagnosis of IDA is challenging when there is coexisting inflammation, as ferritin levels can rise above the normal ranges, even in the presence of iron deficiency. In this case, further tests are required in order to obtain an accurate diagnosis.
In medical laboratories, levels of serum ferritin are commonly used as a measure or indicator of the total amount of iron that is found within the body. The level of serum ferritin is also routinely measured when diagnosing IDA or anaemia of chronic disease. The reference interval is used as a standard to re-test in laboratories for the normal levels of ferritin in the blood: levels of 30 to 300 ng/ml for men and 10 to 200 ng/ml for women are generally regarded as normal, and levels of serum ferritin below 12 ng/ml indicates depletion of iron reserves (Gkamprela et al., 2017; Wang et al., 2010).
Applications of Ferritin
Other than for diagnostic applications, ferritin may also be utilised in the field of materials science as a precursor in the production of iron nanoparticles which are used to grow carbon nanotubes.
These carbon nanotubes have an array of applications in sectors such as nanotechnology, optics, drug delivery, and even as antennas for radios or electromagnetic tools (Anzar et al., 2020).
Ferritin has also been used to create cavities that can act as reaction chambers where metal nanoparticle production can take place. These chambers are able to act as a covering, which prevents the nanoparticles from coagulating. Such ferritin cages, when genetically modified and combined with cell-specific targeting peptides, may also be useful for the targeting of cancerous cells (Uchida et al., 2006). This is made possible due to its distinctive design of 24 self-assembling subunits, hollow chamber capable of encasing pharmaceuticals, and an outside surface that can be genetically and chemically altered for extra functionality.
Ferritin as a Therapeutic
Ferritin nanocages have many advantages over their conventional counterparts. These include their favourable safety profile and high drug loading efficacy. The development of ferritins as drug nanocarriers has quickly increased over the past ten years. Ferritin nanocarriers have the potential to transform therapeutic pharmaceuticals and diagnostic substances into multifunctional nanomedicines as a result of these advantages (Song et al., 2021).
Ferritin in Influenza
In a structural analysis investigation of ferritin carried out by Kanekiyo et al. (2013), researchers found that a heterologous protein, such as influenza virus ‘HA’, may be inserted into ferritin to allow it to take on the physiologically significant trimeric viral spike. Ferritin nanoparticles are made up of 24 subunits organised with octahedral symmetry around a hollow interior, forming a roughly spherical particle. Researchers fused HA to the ferritin nanoparticle, and inserted it into the interface of adjacent subunits, where it generated eight trimeric viral spikes on its surface. The hemagglutination inhibition (HAI) antibody assay was used to titrate the antibody response to an HA infection, and immunisation with this influenza nanoparticle vaccine resulted in HAI titers that were >10 times higher than those from the approved inactivated vaccine. It also induced neutralising antibodies to the stem and the receptor binding site on the head. The stem and the receptor binding site are two highly conserved and susceptible HA sites that are targets of universal vaccinations (Kanekiyo et al., 2013).
.png)
Image 7 - Ferritin as a Therapeutic for Influenza
Ferritin as a therapeutic for cancer
Cancer is the third leading cause of death worldwide, and the global burden of cancer is increasing. The primary treatment option for most solid tumours is surgery, followed by adjuvant chemotherapy to prevent metastasis (World Health Organization, 2022).
However, chemotherapy is the first clinical approach for recurrent cancer. The primary benefit of chemotherapy is that it has a systemic effect on both primary and metastatic tumours.
Doxorubicin (DOX) is one of the most widely used chemotherapeutics in the treatment of solid tumours, but its efficacy in clinical practise is limited by the development of resistance and the occurrence of severe side effects, such as cardiotoxicity and myelosuppression, caused by high dosages. DOX is subjected to the effects of multi-drug resistance mechanisms (MDR), which remove the drug from the cytoplasm, preventing it from exerting its cytotoxic action (Torti et al., 2018).
.png)
Image 8 - Ferritin carrying Doxorubicin
As a result, raising the DOX therapeutic index is critical in cancer research. Recent efforts have resulted in novel DOX delivery strategies, such as the use of liposomes or inorganic nanoparticles, with the goal of reducing drug-related toxicity and evading MDR mechanisms. Apoferritin nano-shells have been proposed as a potentially useful and versatile solution (Torti et al., 2018).
Mammalian ferritins are made up of two types of self-assembling subunits known as H (heavy) and L (light) chains. The H chain contains a catalytic ferroxidase site that catalyses the oxidation of Fe(II) to Fe(III), whereas the L chain participates in the iron nucleation process. Furthermore, because the subunits can be disassembled at an acidic pH and reassembled by bringing the pH back to neutrality in a shape-memory fashion, apoferritin can be used to encapsulate a variety of organic molecules, making it an intriguing scaffold for the development of a biocompatible drug delivery system. In humans, ferritin is specifically recognised by the transferrin receptor 1 (TfR1), which is found overexpressed in many types of tumour cells but not in healthy tissues. An RGD-modified apoferritin cage was recently shown to improve DOX delivery in the cytoplasm of a glioblastoma cell line (Conrad & Pratt, 2019).
In conclusion, ferritin is a complex protein that serves a variety of purposes in the body. It is crucial for immune system control, antioxidant defence, and the storage and regulation of iron. A variety of diseases and conditions, including iron excess and iron deficiency anaemia, can be indicated by abnormal ferritin levels. Clinicians can use a quick blood test to evaluate ferritin levels, which is a crucial diagnostic tool. Ferritin's critical role in preserving our health and wellbeing may be better appreciated if we are aware of its significance and function in the body.
Bibliography
Alkattan, A., Alabdulkareem, K., Kamel, A., Abdelseed, H., Almutairi, Y., & Alsalameen, E. (2021). Correlation betwefen Micronutrient plasma concentration and disease severity in COVID-19 patients. Alexandria Journal of Medicine, 57(1), 21-27. https://10.1080/20905068.2020.1870788
Anzar, N., Hasan, R., Tyagi, M., Yadav, N., & Narang, J. (2020). Carbon nanotube - A review on Synthesis, Properties and plethora of applications in the field of biomedical science. Sensors International, 1, 100003. https://10.1016/j.sintl.2020.100003
Chen, G., Wu, D., Guo, W., Cao, Y., Huang, D., Wang, H., Wang, T., Zhang, X., Chen, H., Yu, H., Zhang, X., Zhang, M., Wu, S., Song, J., Chen, T., Han, M., Li, S., Luo, X., Zhao, J., & Ning, Q. (2020). Clinical and immunological features of severe and moderate coronavirus disease 2019. The Journal of Clinical Investigation, 130(5), 2620-2629. https://10.1172/JCI137244
Cotler, S. J., Bronner, M. P., Press, R. D., Carlson, T. H., Perkins, J. D., Emond, M. J., & Kowdley, K. V. (1998). End-stage liver disease without hemochromatosis associated with elevated hepatic iron index. Journal of Hepatology, 29(2), 257-262. https://10.1016/S0168-8278(98)80011-1
Eng, S. C., Taylor, S. L., Reyes, V., Raaka, S., Berger, J., & Kowdley, K. V. (2005). Hepatic iron overload in alcoholic end-stage liver disease is associated with iron deposition in other organs in the absence of HFE-1 hemochromatosis. Liver International, 25(3), 513-517. https://10.1111/j.1478-3231.2005.01004.x
Gkamprela, E., Deutsch, M., & Pectasides, D. (2017). Iron deficiency anemia in chronic liver disease: etiopathogenesis, diagnosis and treatment. Annals of Gastroenterology, 30(4), 405-413. https://10.20524/aog.2017.0152
Melo, A. K. G., Milby, K. M., Caparroz, A. L. M. A., Pinto, A. C. P. N., Santos, R. R. P., Rocha, A. P., Ferreira, G. A., Souza, V. A., Valadares, L. D. A., Vieira, R. M. R. A., Pileggi, G. S., & Trevisani, V. F. M. (2021). Biomarkers of cytokine storm as red flags for severe and fatal COVID-19 cases: A living systematic review and meta-analysis. PloS One, 16(6), e0253894. https://10.1371/journal.pone.0253894
Oikonomou, T., Goulis, I., Soulaidopoulos, S., Karasmani, A., Doumtsis, P., Tsioni, K., Mandala, E., Akriviadis, E., & Cholongitas, E. (2017). High serum ferritin is associated with worse outcome of patients with decompensated cirrhosis. Annals of Gastroenterology : Quarterly Publication of the Hellenic Society of Gastroenterology, 30(2), 217-224. https://10.20524/aog.2016.0112
Theil, E. C. (2012). Ferritin protein nanocages—the story. Nanotechnology Perceptions, 8(1), 7-16. https://10.4024/N03TH12A.ntp.08.01
Tornai, D., Antal-Szalmas, P., Tornai, T., Papp, M., Tornai, I., Sipeki, N., Janka, T., Balogh, B., & Vitalis, Z. (2021). Abnormal ferritin levels predict development of poor outcomes in cirrhotic outpatients: a cohort study. BMC Gastroenterology, 21(1), 94. https://10.1186/s12876-021-01669-w
Uchida, M., Flenniken, M. L., Allen, M., Willits, D. A., Crowley, B. E., Brumfield, S., Willis, A. F., Jackiw, L., Jutila, M., Young, M. J., & Douglas, T. (2006). Targeting of Cancer Cells with Ferrimagnetic Ferritin Cage Nanoparticles. Journal of the American Chemical Society, 128(51), 16626-16633. https://10.1021/ja0655690
Wang, W., Knovich, M. A., Coffman, L. G., Torti, F. M., & Torti, S. V. (2010). Serum ferritin: Past, present and future. Biochimica Et Biophysica Acta, 1800(8), 760-769. https://10.1016/j.bbagen.2010.03.011
Wang, W., Huang, B., Zhu, Y., Tan, W., & Zhu, M. (2021). Ferritin nanoparticle-based SARS-CoV-2 RBD vaccine induces a persistent antibody response and long-term memory in mice. Cellular & Molecular Immunology, 18(3), 749-751. https://10.1038/s41423-021-00643-6
Huang, X., Chisholm, J., Zhuang, J., Xiao, Y., Duncan, G., Chen, X., Suk, J. S., & Hanes, J. (2017). Protein nanocages that penetrate airway mucus and tumor tissue. Proceedings of the National Academy of Sciences - PNAS, 114(32), E6595-E6602. 10.1073/pnas.1705407114
Kanekiyo, M., Wei, C., Yassine, H. M., McTamney, P. M., Boyington, J. C., Whittle, J. R. R., Rao, S. S., Kong, W., Wang, L., & Nabel, G. J. (2013). Self-Assembling Influenza Nanoparticle Vaccines Elicit Broadly Neutralizing H1N1 Antibodies. Nature (London), 499(7456), 102-106. 10.1038/nature12202
Alunno, A., Carubbi, F., & Rodríguez-Carrio, J. (2020). Storm, typhoon, cyclone or hurricane in patients with COVID-19? Beware of the same storm that has a different origin. RMD Open, 6(1), e001295. https://doi.org/10.1136/rmdopen-2020-001295
Arosio, P., Elia, L., & Poli, M. (2017). Ferritin, cellular iron storage and regulation. IUBMB Life, 69(6), 414–422. https://doi.org/10.1002/iub.1621
Arosio, P., Ingrassia, R., & Cavadini, P. (2009). Ferritins: A family of molecules for iron storage, antioxidation and more. Biochimica et Biophysica Acta (BBA) - General Subjects, 1790(7), 589–599. https://doi.org/10.1016/j.bbagen.2008.09.004
Colafrancesco, S., Alessandri, C., Conti, F., & Priori, R. (2020). COVID-19 gone bad: A new character in the spectrum of the hyperferritinemic syndrome? Autoimmunity Reviews, 19(7), 102573. https://doi.org/10.1016/j.autrev.2020.102573
Conrad, M., & Pratt, D. A. (2019). The chemical basis of ferroptosis. Nature Chemical Biology, 15(12), 1137–1147. https://doi.org/10.1038/s41589-019-0408-1
Dixon, S. J., Lemberg, K. M., Lamprecht, M. R., Skouta, R., Zaitsev, E. M., Gleason, C. E., Patel, D. N., Bauer, A. J., Cantley, A. M., Yang, W. S., Morrison, B., & Stockwell, B. R. (2012). Ferroptosis: an iron-dependent form of nonapoptotic cell death. Cell, 149(5), 1060–1072. https://doi.org/10.1016/j.cell.2012.03.042
Guan, W., Ni, Z., Hu, Y., Liang, W., Ou, C., He, J., Liu, L., Shan, H., Lei, C., Hui, D. S. C., Du, B., Li, L., Zeng, G., Yuen, K.-Y., Chen, R., Tang, C., Wang, T., Chen, P., Xiang, J., & Li, S. (2020). Clinical Characteristics of Coronavirus Disease 2019 in China. New England Journal of Medicine, 382(18). https://doi.org/10.1056/nejmoa2002032
Harigae, H., Hino, K., & Toyokuni, S. (2019). Iron as Soul of Life on Earth Revisited: From Chemical Reaction, Ferroptosis to Therapeutics. Free Radical Biology and Medicine, 133, 1–2. https://doi.org/10.1016/j.freeradbiomed.2019.01.042
He, D., Hughes, S., Vanden-Hehir, S., Georgiev, A., Altenbach, K., Tarrant, E., Mackay, C. L., Waldron, K. J., Clarke, D. J., & Marles-Wright, J. (2016). Structural characterization of encapsulated ferritin provides insight into iron storage in bacterial nanocompartments. ELife, 5, e18972. https://doi.org/10.7554/eLife.18972
Huang, I., Pranata, R., Lim, M. A., Oehadian, A., & Alisjahbana, B. (2020). C-reactive protein, procalcitonin, D-dimer, and ferritin in severe coronavirus disease-2019: a meta-analysis. Therapeutic Advances in Respiratory Disease, 14, 175346662093717. https://doi.org/10.1177/1753466620937175
Jian, N., Dowle, M., Horniblow, R. D., Tselepis, C., & Palmer, R. E. (2016). Morphology of the ferritin iron core by aberration corrected scanning transmission electron microscopy. Nanotechnology, 27(46), 46LT02. https://doi.org/10.1088/0957-4484/27/46/46lt02
Lundin, D., Poole, A. M., Sjöberg, B.-M., & Högbom, M. (2012). Use of structural phylogenetic networks for classification of the ferritin-like superfamily. The Journal of Biological Chemistry, 287(24), 20565–20575. https://doi.org/10.1074/jbc.M112.367458
Ng, S. S., Kavanagh, K. L., McDonough, M. A., Butler, D., Pilka, E. S., Lienard, B. M. R., Bray, J. E., Savitsky, P., Gileadi, O., von Delft, F., Rose, N. R., Offer, J., Scheinost, J. C., Borowski, T., Sundstrom, M., Schofield, C. J., & Oppermann, U. (2007). Crystal structures of histone demethylase JMJD2A reveal basis for substrate specificity. Nature, 448(7149), 87–91. https://doi.org/10.1038/nature05971
Ruscitti, P., Berardicurti, O., Barile, A., Cipriani, P., Shoenfeld, Y., Iagnocco, A., & Giacomelli, R. (2020). Severe COVID-19 and related hyperferritinaemia: more than an innocent bystander? Annals of the Rheumatic Diseases, 79(11), 1515–1516. https://doi.org/10.1136/annrheumdis-2020-217618
Siddiqi, H. K., & Mehra, M. R. (2020). COVID-19 Illness in Native and Immunosuppressed States: A Clinical-Therapeutic Staging Proposal. The Journal of Heart and Lung Transplantation, 39(5). https://doi.org/10.1016/j.healun.2020.03.012
Theil, E. C. (2013). Ferritin: The Protein Nanocage and Iron Biomineral in Health and in Disease. Inorganic Chemistry, 52(21). https://doi.org/10.1021/ic400484n
Torti, S. V., Manz, D. H., Paul, B. T., Blanchette-Farra, N., & Torti, F. M. (2018). Iron and Cancer. Annual Review of Nutrition, 38(1), 97–125. https://doi.org/10.1146/annurev-nutr-082117-051732
Wang, D., Hu, B., Hu, C., Zhu, F., Liu, X., Zhang, J., Wang, B., Xiang, H., Cheng, Z., Xiong, Y., Zhao, Y., Li, Y., Wang, X., & Peng, Z. (2020). Clinical Characteristics of 138 Hospitalized Patients with 2019 Novel Coronavirus–Infected Pneumonia in Wuhan, China. JAMA, 323(11), 1061–1069. https://doi.org/10.1001/jama.2020.1585
Bellini, M., Mazzucchelli, S., Galbiati, E., Sommaruga, S., Fiandra, L., Truffi, M., Rizzuto, M. A., Colombo, M., Tortora, P., Corsi, F., & Prosperi, D. (2014). Protein nanocages for self-triggered nuclear delivery of DNA-targeted chemotherapeutics in Cancer Cells. Journal of Controlled Release, 196, 184–196. https://doi.org/10.1016/j.jconrel.2014.10.002
World Health Organization. (2022, February 3). Cancer. World Health Organization. https://www.who.int/news-room/fact-sheets/detail/cancer